A look into the neurophysiology behind the runner's high
By Leanne Fogg

Running 13 miles for fun is often thought of as crazy, but for most long distance athletes it is simply another workout that comes with a blissful reward: the runner’s high. Toward the end of a long run, I find myself easily holding a fast pace. My body maintains a smooth rhythm, executing perfect biomechanics as I float along the trail. When in company, I can share this cadence and continue for miles in silence, enjoying the sounds of our synchronized steps. I can find myself in a state of mindlessness, escaping the weight of the stressors of life. This state of tranquility and harmony is referred to as a “runner’s high,” and results after intense exercise.
Reported effects of the runner’s high mirrored the
marijuana-induced state many are familiar with. Yes, runners are claiming they can
get baked without smoking.
A wave of skepticism triggered the investigation of the
runner’s high hypothesis – can exercise alter the neurophysiology of an
athlete, leading to pain-relieving and calming effects?
Before we dive into the exercise-induced high investigation,
it is helpful to understand the neurophysiological effects marijuana has on the
body.
The effects of marijuana are mainly attributed
to Δ9-tetrahydrocannabinol
(THC) – the primary psychoactive component in the cannabis plant. THC alters
the cell-cell communication in the nervous system. A signaling presynaptic neuron will release neurotransmitters, which will bind to their
corresponding receptors on the surface of a target postsynaptic neuron. Receptors
contain highly specific binding domains that only particular molecules can fit
into and elicit the downstream response. Although receptors are designed for
specificity, exogenous molecules, like THC, can hijack systems and trigger
alterations in many biological processes.
Scientist first discovered these receptors as being sensitive to
THC, and appropriately named them the cannabinoid receptors. Cannabinoid
receptors come in two flavors: type 1 (CB1R) and type 2 (CB2R). CB1 receptors
are primarily located in the central nervous system, as it is one of the most
prevalent G protein-coupled receptors in the brain. CB2 receptors are found in
tissues throughout the body, and are involved in mediating the immune response.
 |
The cannabinoid receptors bind endocannabioids (endogenous
neurotransmitters) including arachidonoylethanolamine (anandamide or AEA) and γ-aminobutyric
acid (2-AE). Anandamide, meaning “blissful amide,” is the main agonist (effector molecule) of CB1 receptors, leading to alterations in appetite,
mood, and pain sensation.
Although receptors are highly specific, CB1 receptor activation can result through the binding of an endocannabinoid or THC. CB1 receptor binding occurs through the recognition of a hydrocarbon chain motif that AEA, 2-AE, and THC all share.
 |
| CB1 receptor agonists (highlighted: binding motifs) |
Endocannabinoid receptors are a type of G protein – they are protein receptors that span the membrane of a cell and trigger a signal transduction pathway upon
binding of a specific molecule. CB1 receptors are located at the axon terminals
of a presynaptic neuron (the “end” of a signaling neuron). Binding to the CB1 receptor will
prohibit the release of neurotransmitters, inhibiting the
signal from being passed to the postsynaptic neuron (the target neuron will not
receive the signal). This process
naturally occurs in our bodies and is known as depolarization-induced-suppression
inhibition (DSI).
 |
| Depolarization-induced-suppression inhibition (DSI) induced through binding of CB1 receptor |
Whoa there... DSI? Let’s break this down:
Depolarization-induced: This effect is triggered upon
depolarization of a neuron (when an action potential arrives, the cell
depolarizes, becoming more positively charged on the inside).
Suppression: CB1 receptors are located on GABAergic neurons
(neurons that release the neurotransmitter γ-aminobutyric acid, or GABA). GABA
is an inhibitory neurotransmitter, suppressing the target cell from depolarization.
Inhibition: GABA neurotransmitter release is inhibited through the
activation of CB1 receptors.
All together now: When postsynaptic neurons become depolarized, endocannibinoids
are released and travel retrograde to CB1 receptors on a GABAergic neuron.
Activation of the CB1 receptor then leads to the inhibition of GABA release.
CB1 receptor activation leads to the inhibition of adenylyl
cyclase (the enzyme that synthesizes cAMP), and subsequent decrease in calcium
concentrations. Low amounts of calcium prevent GABA-containing vesicles from
fusing out of the cell. DSI alters neuronal communication, changing the way we perceive pain and stress.
So how is this all connected to the runner’s high
hypothesis? It has been demonstrated that endocannabinoid system (ECS) activation
can be induced upon exercise in humans.
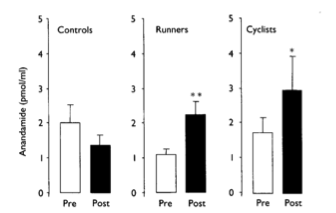 |
| Circulating AEA levels in human subjects were significantly higher only after intense running or cycling (Sparling, 2003). |
A comparison between cursorial (an animal adapted for running) and non-cursorial mammals revealed elevated AEA levels after exercise in dogs and humans while no significant increase in ferrets.
 |
| A) Plasma AEA levels before (white) and after (black) high intensity running; B) plasma AEA levels before (white) and after (black) walking, (Raichlen, 2011). |
This study sheds light on possible evolutionary benefits of ECS activation. Allowing animals that have pushed themselves to their limit to enter a euphoric state might allow them to better survive - maybe running even farther than they anticipated to catch prey, or even running from a predator. ECS activation could be a result of a mechanism that promotes endurance in running mammals.
Circulating AEA has been demonstrated to correlate with the subsequent binding to CB1 receptors in specific regions throughout the brain, including the frontolimbic brain areas associated with mood. Euphoric levels reported by subjects correlated with CB1 receptor binding in the brain.
Activation of ESC has also provided evidence of reducing
pain sensitivity and sedation, further supporting the runner's high hypothesis.
 |
| fMRI scans before and after exercise show a reduction in pain processing in the brain (Scheef, 2012). |
Exercise increases AEA levels, which in turn act on CB1 receptors, leading to DSI. It is through this mechanism that we can enter euphoric states - whether that be smoking dope or going for a long run on the central coast... I prefer the latter.
Stay tuned for more on the endocannabinoid system and the promising future in the treatment of breast cancer, depression, and obesity.
References:
Alger, B.E., Endocannabinoid at the synapse a decade after the dies mirabilis (29 March 2001): what we still do not know, The Journal of Physiology 590(10):2203-2212.
Boecker, H., T. Sprenger, M.E. Spilker, G. Henriksen, M. Koppenhoefer, K.J. Wagner, M. Valet, A. Berthele, T.R. Tolle. 2008. The runner's high: opioidergic mechanism in the human brain, Cerebral Cortex 18:2523-2531.
Raichlen, D. A., A.D. Foster, G.L. Gerdeman, A. Seiller, A. Giuffrida. 2011. Wired to run: exercise-induced endocannabinoid signaling in humans and cursorial mammals with implications for the 'runner's high, The Journal of Experimental Biology 215:1331-1336.
Scheef, L., J. Jankowski, M. Daamen, G. Weyer, M. Klingenberg, J. Renner, S. Mueckter, B. Schurmann, F. Musshoff, M. Wagner, H.H. Schild, A. Zimmer, H. Boecker, An fMRI study on the acute effect of exercise on pain processing in trained athletes, International Association for the Study of Pain 153:1702-1714.
Sparling, P.B., A. Giuffrida, D. Piomelli, L. Rosskopf, A. Dietrich. 2003. Exercise activates the endocannabinoid system, NeuroReport 14(17):2209-2211.
Terry, G.E., J. Liow, S.S. Zoghbi, J. Hirvonen, A.G. Farris, A. Lerner, J.T. Tauscher, J.M. Schaus, L. Phebus, C.C. Felder, C.L. Morse, J.S. Hong, V.W. Pike, C. Halldin, R.B. Innis, Quantitation of cannabinoid CB1 receptors in healthy human brain using positron emission tomography and an inverse agonist radioligand, Neuroimage 48(2):362-370.
I'm assuming you can get this "high" from other types of endurance/aerobic exercise?
ReplyDeleteRunning is one of the most common forms of exercise used in these studies, as it is good way to keep your heart rate elevated for long periods of time. Sparling observed an increase in plasma AEA levels in cyclists after two hours of biking. I believe it is not the specific locomotion of running that activated the ECS, but the body's response to high intensity exercise.
ReplyDeleteGreat topic! I have long wondered why sometimes when running I feel so good that I run much longer than I intend to -- this blog explains the "runner's high" quite well. Awesome job
ReplyDeleteSo this must be why stoners are so lazy...it's so much easier to just light up a doobie. Though obviously not as healthy or rewarding. Good job Leanne!
ReplyDeleteI was wondering the other day as I was running with my dog and experiencing this wonderful "high," if she.... as she lagged behind with her tongue practically on the ground... was, as well. Then I thought about all those stories of "getting your [poor] dog/cat high" and wondered if they, or other mammals in general, have the same cannabanoid receptor system.... It would sure be nice for Rue to enjoy her run as much as I do!!
ReplyDeleteHealing from HIV-AIDS, i never though dr.camala who could ever get my HIV-AIDS cured with his healing spell, i have tried almost everything but i couldn't find any solution on my disease, despite all these happening to me, i always spend alot to buy a HIV drugs from hospital and taking some several medications but no relieve, until one day i was just browsing on the internet when i come across a great post of !Michelle! who truly said that she was been diagnose with HIV and was healed that very week through the help of these great powerful healing spell doctor, sometime i really wonder why people called him Papa camala, i never knew it was all because of the great and perfect work that he has been doing that is causing all this. so i quickly contacted him, and he ask me some few questions and he said a thing i will never forget that anyone who contacted him is ! always getting his or her healing in just 6 hours after doing all he ask you, so i was amazed all the time i heard that from him, so i did all things only to see that at the very day which he said i will be healed, all the strength that has left me before rush back and i becomes very strong and healthy, this disease almost kills my life all because of me, so i will to hospital to give the final test to the disease and the doctor said i am HIV negative, i am very amazed and happy about the healing dr.camala gave to me from the ancient part of Africa, you can email him now for your own healing too at: dr.camalahivadscure@yahoo.com or WhatsApp him +2349055637784 thank you sir for healing me from HIV, i am Doris Carter.
ReplyDeleteI Want To Appreciate Dr.OYAGU for his great deeds, I Was Diagnosed With type 2 Herpes Virus Last year,And i Was Looking For Solution To Be Cured Luckily I Saw Testimonies On How Dr.OYAGU Cure Herpes Virus I Decided To Contact Dr.OYAGU I Contacted Him He Prepared A Herbal Medicine Portion And Sent It To Me,I Started The Herbal Medicine For My Health.He Gave Me Step By Step Instructions On How To Apply It, When I Applied It As Instructed, I Was Cured Of This Deadly Herpes Within 2 weeks, I Am Now Herpes Negative.My Brother And Sister I No That There Are So Many People That Have The Same Herpes Virus Please contact Dr OYAGU To Help You Too,And Help Me To Thank Dr.OYAGU For Cure Me, I’m Cured By Dr. OYAGU Herbal Medicine,His Contact Email:oyaguherbalhome@gmail.com
ReplyDeleteOr Cell Whatsapp Number +2348101755322 thank you